Brachiosaurus
Brachiosaurus altithorax
"Arm lizard with high thorax"
Sobre esta espécie
Brachiosaurus altithorax is one of the largest dinosaurs ever discovered in North America. It lived during the Late Jurassic, approximately 154 to 143 million years ago, on the alluvial plains of the Morrison Formation. It is distinguished by forelimbs longer than its hindlimbs, an extremely long neck, and a high thorax, which allowed it to reach vegetation more than 9 meters above the ground. Originally described by Elmer Riggs in 1903, it was for decades confused with its African relative Giraffatitan brancai, whose definitive taxonomic separation only occurred in 2009. With more than 26 distinct skeletal characteristics, it represents an exclusively North American genus.
Geological formation & environment
The Morrison Formation is a sequence of Late Jurassic sedimentary rocks (Kimmeridgian-Tithonian, ~155–148 Ma) extending across eight American states, from Montana to New Mexico. It represents one of the largest and most diverse dinosaur faunas in the world. The original environment was semi-arid alluvial plains, with seasonal rivers, ponds, and riparian vegetation dominated by conifers, ginkgos, and cycads. The Brushy Basin Member, where the Brachiosaurus altithorax holotype was found near Fruita, Colorado, is particularly rich in vertebrate fóssils. The formation also preserves Allosaurus, Diplodocus, Camarasaurus, Apatosaurus, Stegosaurus, and dozens of other species.
Image gallery







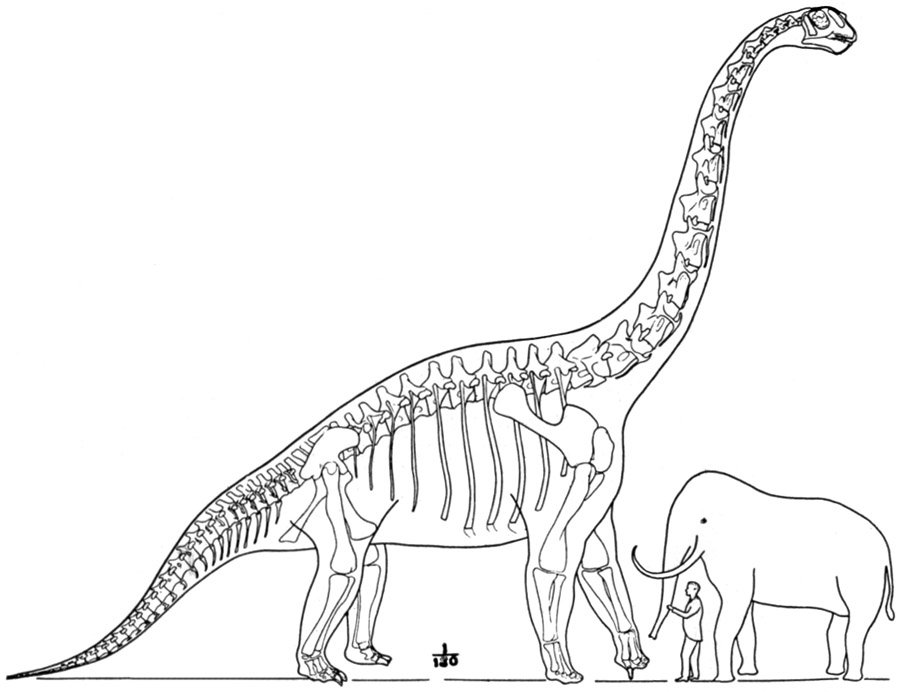



Digital reconstruction of Brachiosaurus altithorax by Dmitry Bogdanov (2007), based on Scott Hartman's skeletal proportions. This is one of the most widely used representations in scientific publications.
Dmitry Bogdanov / Domínio Público
Ecology and behavior
Habitat
Brachiosaurus altithorax inhabited the alluvial plains of the Morrison Formation during the Late Jurassic, 154 to 143 million years ago. The climate was semi-arid, with a prolonged dry season and irregular seasonal precipitation, similar to a modern savanna. Vegetation was dominated by arboreal conifers, ginkgos, cycads, ferns, and horsetails, with no angiosperms. Seasonal rivers and flood plains provided access to water and dense vegetation. Brachiosaurus coexisted with Allosaurus, Ceratosaurus, Diplodocus, Camarasaurus, Apatosaurus, and Stegosaurus in the same ecosystem.
Feeding
With a neck up to 9 meters raised vertically, Brachiosaurus reached canopy vegetation inaccessible to other Morrison Formation sauropods. Its wide, spatulate teeth were adapted for shearing conifer branches and ginkgo canopies, not for processing ground-level vegetation. The position of the skull atop the neck suggested continuous 'canopy browsing,' exploiting vertical strata above 5 meters. To meet the metabolism of a 50–60 tonne animal, it needed to consume hundreds of kilograms of vegetation per day, directly shaping the structure of local vegetation.
Behavior and senses
The fóssil record of Brachiosaurus altithorax is too fragmentary for direct behavioral inferences. Based on sauropods in general, they likely lived in groups or herds, reducing predation risk from Allosaurus. Locomotion was obligately quadrupedal, without the ability to rear on hindlimbs as older reconstructions proposed. Morrison Formation sauropod trackways show a 'manus-only' pattern consistent with Brachiosaurus biomechanics (longer forelimbs). There is no evidence of parental care or specific reproductive behavior for the species.
Physiology and growth
Extensive vertebral pneumatization in Brachiosaurus evidences avian-style air sacs, indicating highly efficient unidirectional pulmonary ventilation. This respiratory physiology suggests elevated metabolism, incompatible with ectothermy. The blood pressure needed to pump blood to the skull at 9 meters height — estimated above 700 mmHg — would imply a high-performance heart. Rapid growth inferred from bone histology of related sauropods is consistent with endothermy. Estimated weight between 28 and 58 tonnes depending on estimation method. Body temperature likely regulated by a combination of internal metabolism and thermal inertia (gigantothermy).
Paleogeography
Continental configuration

Ron Blakey · CC BY 3.0 · Jurassic, ~90 Ma
Fóssil sites
Maltese, Tschopp, Holwerda, Burnham, PeerJ (2018) · CC BY 4.0
During the Kimmeridgiano-Titoniano (~154.8–143.1 Ma), Brachiosaurus altithorax inhabited the fragmenting Pangea. North America and Europe were still close, and the North Atlantic was just beginning to open. Climate was warm and humid globally, with no polar ice caps.
Inventário de Ossos
The holotype FMNH P25107 preserves the right humerus, right fêmur, right ilium, right coracoid, sacrum, last seven thoracic and two caudal vertebrae, and partial ribs. No skull was found associated with the holotype. Additional specimens from Utah, Wyoming, and Oklahoma have slightly expanded the inventory, but Brachiosaurus altithorax remains a fragmentary taxon.
Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
Brachiosaurus altithorax, the largest known dinosaur
Riggs, E.S. · American Journal of Science
Founding paper of all Brachiosaurus paleontology. Elmer Riggs describes specimen FMNH P25107, collected from the Morrison Formation near Fruita, Colorado in 1900, establishing the genus Brachiosaurus and species altithorax. The genus name refers to the disproportionately long arms: the humerus (204 cm) and fêmur (203 cm) are nearly identical in length, unique among sauropods. The epithet 'altithorax' describes the elevated thorax. Riggs argues, pioneering for the time, that sauropods were terrestrial animals rather than aquatic amphibians as the dominant view held. The holotype preserves the right humerus, fêmur, ilium, coracoid, sacrum, thoracic and caudal vertebrae. Riggs estimates the animal would be the largest known dinosaur. This paper is the mandatory starting point for any study of the species.

Structure and relationships of opisthocoelian dinosaurs. Part II. The Brachiosauridae
Riggs, E.S. · Geológical Series of the Field Columbian Museum
Supplementary osteological monograph to the 1903 original description. Riggs deepens the anatômical analysis of Brachiosaurus altithorax, providing detailed description of the morphology of opisthocoelous vertebrae (with convex posterior face), limb bones, and pelvic girdle. He formally establishes the family Brachiosauridae, positioning Brachiosaurus in relation to other sauropods then known. Analysis of the dorsal vertebrae reveals extensive pleurocels (lateral cavities), correctly interpreted as pneumatic chambers connected to the respiratory system. The work includes detailed measurements of each preserved bone element and high-quality illustrated plates. Riggs consolidates the terrestrial hypothesis for the animal, arguing that limb structure is incompatible with aquatic habits. This monograph remains the primary anatômical description of the holotype.
The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world's largest dinosaurs
Paul, G.S. · Hunteria
Pioneering work that first formally questions the taxonomic unity of Brachiosaurus altithorax and the African material then called B. brancai. Paul identifies substantial morphological differences between the two fóssil sets: different proportions of trunk, neck, and limbs, plus distinct cranial architectures. He proposes that the African material should be classified as a separate subgenus, which he names Giraffatitan. Includes body mass estimates for both forms and comparison with other large dinosaurs of the time. Although Paul proposes only subgeneric separation, he plants the debate that will culminate 21 years later in full generic separation by Taylor (2009). The paper also provides body reconstructions based on skeletal proportions, without the cladistic rigor of modern paleontology.

A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai
Taylor, M.P. · Journal of Vertebrate Paleontology
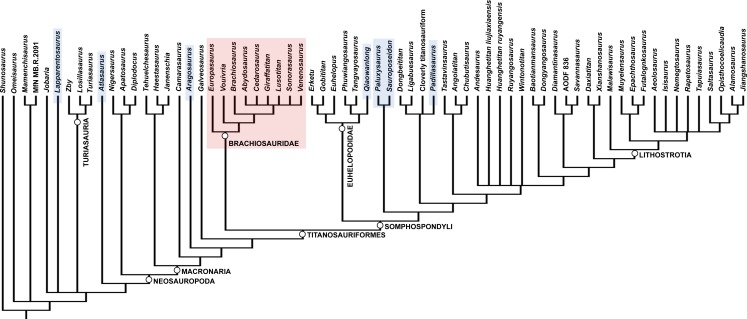
The most important paper in the modern history of Brachiosaurus. Michael Taylor performs exhaustive comparative analysis of North American Brachiosaurus altithorax fóssils and African material then called B. brancai, cataloguing 26 distinct osteological differences: Brachiosaurus has a dorsal vertebral series 23% longer, a tail 20–25% larger, vertebrae with fundamentally different morphology, and limbs with distinct proportions. Taylor concludes the differences are comparable to those separating Diplodocus from Barosaurus, fully justifying two separate genera. The African material is renamed Giraffatitan brancai. The separation has immediate impact: most museums worldwide were displaying what they believed was Brachiosaurus, but were actually Giraffatitan. The paper also provides an emended diagnosis of Brachiosaurus altithorax based on preserved holotype elements, becoming the primary reference for all subsequent studies.
The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae
Mannion, P.D., Allain, R. & Moine, O. · PeerJ
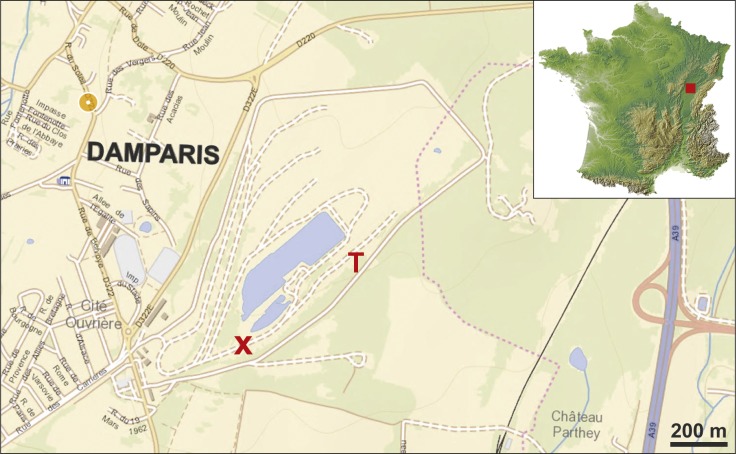
High-impact paper redefining the evolutionary history of Brachiosauridae. Mannion, Allain, and Moine describe Vouivria damparisensis, a skeleton discovered in France in 1934 previously called the 'French Bothriospondylus', now identified as the earliest known titanosauriform (~160 Ma, middle-late Oxfordian). Phylogenetic analysis of 416 characters across 77 taxa firmly positions Brachiosaurus altithorax within Brachiosauridae, in a clade including Vouivria, Lusotitan, Europasaurus, and Abydosaurus. Results indicate Brachiosauridae was composed exclusively of Late Jurassic forms, with the clade becoming globally extinct in the earliest Late Cretáceous. The work clarifies the biogeography of the group, showing diversification occurred mainly in Laurasia. For Brachiosaurus altithorax, the study confirms its phylogenetic position and importance as a reference for understanding titanosauriform evolution.
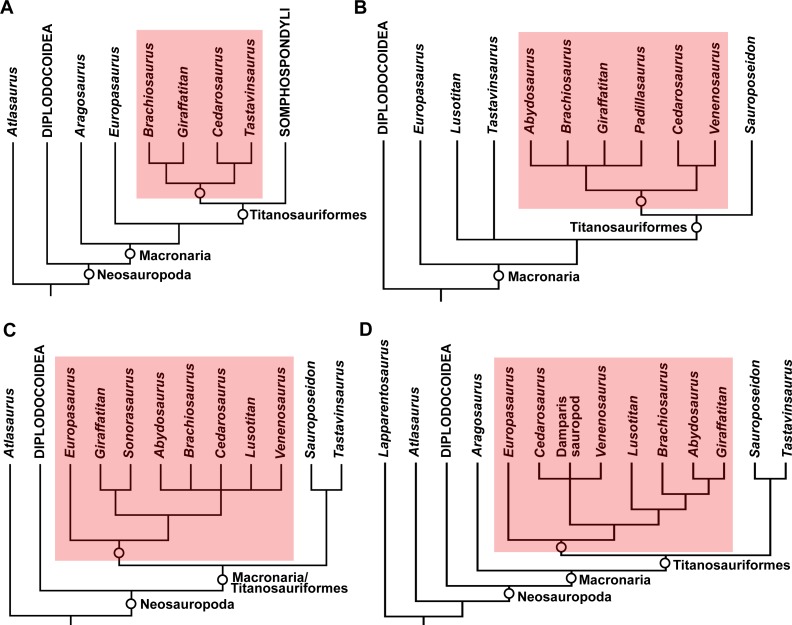
A new Early Cretáceous brachiosaurid (Dinosauria, Neosauropoda) from northwestern Gondwana (Villa de Leiva, Colombia)
Carballido, J.L., Pol, D., Parra Ruge, M.L., Padilla Bernal, S., Páramo-Fonseca, M.E. & Etayo-Serna, F. · Journal of Vertebrate Paleontology
Paper expanding the geographic and temporal record of Brachiosauridae beyond North America and Europe. Carballido et al. describe Padillasaurus leivaensis, a brachiosaurid from the Early Cretáceous of Colombia (Aptian, ~130 Ma), making it the most recent and southernmost representative of the family. Phylogenetic analysis includes Brachiosaurus altithorax as a reference taxon and confirms Brachiosauridae monophyly. The authors discuss the biogeographic paradox: how did brachiosaurids reach Gondwana if the group was predominantly Laurasian? The study suggests transatlantic dispersal during the Late Jurassic or Early Cretáceous, when the South Atlantic was still a shallow sea. This context broadens understanding of Brachiosauridae diversification and Brachiosaurus altithorax's role as a diagnostic reference for the family.
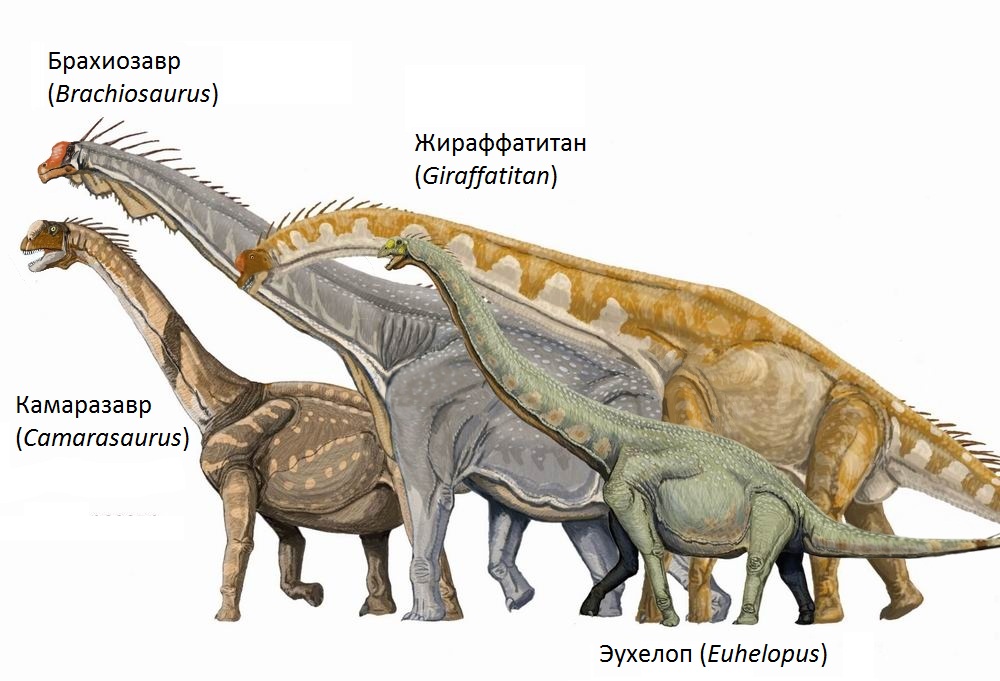
Head and neck posture in sauropod dinosaurs inferred from extant animals
Taylor, M.P., Wedel, M.J. & Naish, D. · Acta Palaeontologica Polonica
Taylor, Wedel, and Naish resolve one of the most debated questions in sauropod biology: in what position did they carry their necks? Using systematic analysis of living long-necked animals (giraffes, swans, eagles, iguanas, crocodilians), the authors demonstrate that in neutral rest these animals hold the neck in the 'natural' position defined by the curvature of cervical articulations. Applying this logic to Brachiosaurus, the neck naturally points upward and forward, not horizontally as proposed by Gunga et al. (1999) and others. The elevated neck posture is consistent with the morphology of Brachiosaurus cervical vertebrae, which show articulations inclined to permit dorsal extension. The paper has direct implications for feeding models and cardiovascular physiology: an erect neck at 9 meters would require extreme blood pressure, additional evidence of endothermic physiology.

Sauropod necks: are they really for heat loss?
Henderson, D.M. · PLOS ONE
Henderson analyzes 16 three-dimensional digital models of sauropods ranging from 639 kg (juvenile Camarasaurus) to 25 tonnes (Brachiosaurus), measuring total body and neck surface áreas to test the hypothesis that long necks functioned as thermal radiators. Results reveal a non-trivial relationship: while total body área shows negative allometry against metabolic rate (as expected), neck área shows positive allometry with exponent 1.17, meaning the neck grows faster than the rest of the body in larger animals. In Brachiosaurus, the neck would represent a disproportionate fraction of body surface, consistent with an additional thermoregulatory function. The study does not dismiss feeding or reproductive functions, but adds the thermoregulatory hypothesis as an evolutionary selective variable. It is one of the first works to quantify the thermal physiology of Brachiosaurus through digital modeling.

Simulating sauropod manus-only trackway formation using finite-element analysis
Falkingham, P.L., Bates, K.T., Margetts, L. & Manning, P.L. · Biology Letters
Falkingham et al. use finite element analysis to simulate how different substrates respond to sauropod weight, comparing Diplodocus and Brachiosaurus. The result has direct implication for Brachiosaurus biology: by having longer forelimbs and a forward-shifted center of mass, Brachiosaurus applied much greater pressure with its manus (forelimbs) than with its pes (hindlimbs). This explains why sauropod trackways often show only forelimb impressions — the pes often left no mark on the substrate because pressure was insufficient. The computational model demonstrates that Brachiosaurus's unique morphology with arms longer than legs is not just an anatômical feature, but has measurable biomechanical consequence in the form of locomotion and the type of trackway left behind. This paper is one of the few to study the locomotor behavior of Brachiosaurus altithorax specifically.

Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs
Wedel, M.J. · Paleobiology
Wedel systematically investigates pleurocels (pneumatic chambers) in sauropod vertebrae and demonstrates these structures are homologous to the air diverticula of modern birds. In Brachiosaurus altithorax, vertebral pneumatization is particularly extensive, reducing axial skeleton mass by up to 20% compared to equivalent solid bone. This has profound implications: with avian-style air sacs, Brachiosaurus likely had highly efficient unidirectional pulmonary ventilation comparable to birds. This respiratory system would be essential to supply the metabolism of a 50–60 tonne animal. The extensive pneumatization of cervical vertebrae would also explain how the 9-meter neck could be raised and sustained without collapsing under its own weight. The paper establishes that Brachiosaurus respiratory physiology was fundamentally different from living reptiles, aligning more with birds than crocodilians.


Redescription of brachiosaurid sauropod dinosaur material from the Upper Jurassic Morrison Formation, USA
D'Emic, M.D. & Carrano, M.T. · The Anatômical Record
D'Emic and Carrano systematically review brachiosaurid material from the Morrison Formation, including specimens previously referred to Brachiosaurus altithorax but never redescribed since Riggs' original works. The authors apply modern cladistic diagnostic criteria to assess which specimens can be confidently attributed to Brachiosaurus altithorax and which represent distinct taxa or indeterminate material. The work identifies additional diagnostic characters not recognized by Taylor (2009) and discusses the completeness of the Brachiosaurus fóssil record in the Morrison Formation. Crucial for understanding how many true Brachiosaurus specimens exist: many fóssils previously attributed to the species are redescribed as indeterminate Brachiosauridae or possible new taxa. The study uses modern tomographic and morphometric analysis techniques to reexamine over-a-century-old material.


A new brachiosaurid sauropod from the Upper Jurassic Morrison Formation of the USA
Maltese, A., Tschopp, E., Holwerda, F. & Burnham, D. · PeerJ
Maltese et al. describe a nearly complete brachiosaurid pes, informally called 'Bigfoot', from the Morrison Formation, representing the most complete hindlimb extremity material ever found for the group in North America. Phylogenetic analysis indicates the specimen belongs to Brachiosauridae and may represent Brachiosaurus altithorax or a related undescribed taxon. The study provides the first detailed description of foot anatomy for a North American brachiosaurid, revealing features such as phalangeal formula and astragalus structure. The distribution map of brachiosaurids in the Morrison Formation identifies ten distinct localities, showing these animals had a broader geographic range than previously recognized. For Brachiosaurus altithorax, the work is relevant for potentially expanding the skeletal inventory with extremity material absent from the holotype.


Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem — a synthesis
Turner, C.E. & Peterson, F. · Sedimentary Geology
Turner and Peterson synthesize decades of geológical, paleobotanical, and paleontológical research to reconstruct the complete Morrison Formation ecosystem in the Late Jurassic, the native environment of Brachiosaurus altithorax. The climate was semi-arid, similar to a modern savanna, with a prolonged dry season and irregular seasonal precipitation. Vegetation was dominated by arboreal conifers, ginkgos, cycads, tree ferns, and horsetails, without angiosperms. In terms of fauna, the Morrison was one of the most biodiverse terrestrial ecosystems of the Mesozoic, with multiple sauropod species (Diplodocus, Camarasaurus, Apatosaurus, Brachiosaurus), theropods (Allosaurus, Ceratosaurus), stegosaurids, ornithopods, and small tetrapods. Brachiosaurus, as the tallest canopy browser in the formation, would have played a key ecológical role in shaping vegetation and nutrient cycling. The paper provides environmental context essential for interpreting Brachiosaurus paleoecology.


Minimum convex hull mass estimations of complete mounted skeletons
Sellers, W.I., Hepworth-Bell, J., Falkingham, P.L., Bates, K.T., Brassey, C.A., Egerton, V.M. & Manning, P.L. · Biology Letters
Sellers et al. develop and validate an objective methodology for estimating dinosaur body mass: three-dimensional laser scanning of mounted skeletons followed by minimum convex hull calculation. The method is tested on 14 large mammal skeletons (elephants, hippopotamuses, rhinoceroses) with known masses, demonstrating consistent underestimation of 21%. Applied to the Giraffatitan skeleton in Berlin's museum (frequently used as a proxy for Brachiosaurus before 2009), the method generates a baseline estimate of 23.2 tonnes, corrected by the 21% underestimation factor to yield ~28.7 tonnes for that specimen. For Brachiosaurus altithorax, which was probably larger, estimates range from 28 to 58 tonnes depending on the method. This paper provides the most rigorous methodology available for estimating the mass of the largest North American Jurassic dinosaur.

First complete sauropod dinosaur skull from the Cretáceous of the Americas and the evolution of sauropod dentition
Chure, D., Britt, B.B., Whitlock, J.A. & Wilson, J.A. · Naturwissenschaften
Chure et al. describe Abydosaurus mcintoshi, an Early Cretáceous brachiosaurid from Utah (104.46 ± 0.95 Ma) with a complete skull, the first for any brachiosaurid from the Americas. This find is fundamental for Brachiosaurus altithorax because it resolves a critical gap: the Brachiosaurus holotype has no skull, and all cranial inference depended on Giraffatitan or isolated fragments. Abydosaurus, as a North American brachiosaurid that 'shares close ancestry with Brachiosaurus,' offers the most legitimate proxy for cranial morphology. Comparison between Abydosaurus and Jurassic specimens shows an evolutionary trend: teeth shrank dramatically over 45 million years, changing from wide and spatulate in Brachiosaurus to narrow and pencil-like in Abydosaurus, suggesting a shift in diet or food processing method. The complete skull reveals nasal, orbital, and temporal structure of brachiosaurids.


Espécimes famosos em museus
FMNH P25107 (Holótipo)
Field Museum of Natural History, Chicago, EUA
The holotype of Brachiosaurus altithorax, discovered on July 4, 1900, near Fruita, Colorado. Preserves the right humerus, fêmur, ilium and coracoid, sacrum, seven distal thoracic vertebrae, and two caudal vertebrae. A skeletal mount based on this specimen with parts supplemented from Giraffatitan is displayed outside the museum, and a replica is at Terminal 1 of O'Hare Airport, Chicago.
USNM 21903 (Úmero de Potter Creek)
Smithsonian National Museum of Natural History, Washington D.C., EUA
The largest known Brachiosaurus altithorax humerus, from Potter Creek, Utah. At 213 cm long, it surpasses the holotype and indicates the animal could have been even larger than Riggs' specimen. The associated ulna (USNM 21903) is displayed at the Smithsonian Museum of Natural History and is one of the few publicly exhibited elements of true Brachiosaurus altithorax.
In cinema and popular culture
Brachiosaurus is one of the most recognizable dinosaurs in popular culture, largely due to the opening scene of Steven Spielberg's Jurassic Park (1993). The moment Sam Neill looks up and sees a Brachiosaurus rearing to eat from a tree canopy is considered one of the most impactful in cinema history. The animal reappeared in Jurassic Park III (2001) as background fauna and had its most emotionally charged scene in Jurassic World: Fallen Kingdom (2018), when a lone specimen perishes in the island's volcanic eruption. The BBC documentary Walking with Dinosaurs (1999) portrayed the animal in the correct Morrison Formation context. Disney's Dinosaur (2000) anthropomorphized the animal into the character Baylene. In all these cases, a persistent scientific inaccuracy remained: before 2009, depictions were based on Berlin's Giraffatitan, not the North American Brachiosaurus altithorax, resulting in incorrect proportions. Even so, Brachiosaurus's cultural role as a symbol of grandeur and peace is unquestionable.

Classificação
Descoberta
Curiosidade
Brachiosaurus altithorax had forelimbs longer than its hindlimbs: the humerus (arm) at 204 cm and fêmur (thigh) at 203 cm are nearly identical in length, the opposite of what occurs in most dinosaurs. That is why the genus is called 'arm lizard': the arms are its most extraordinary feature.