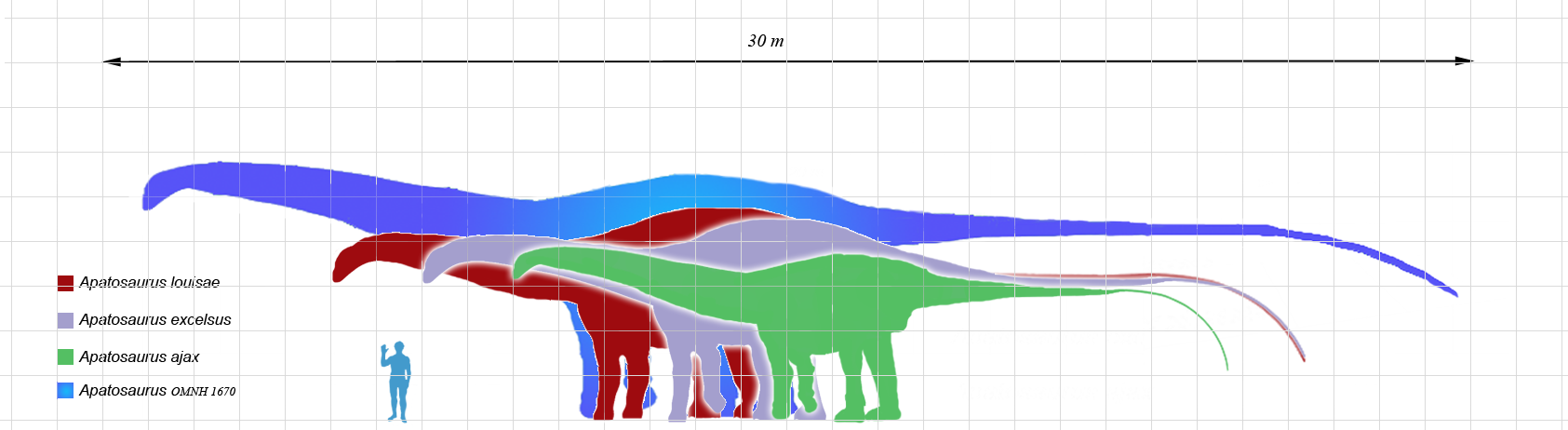
Brontosaurus
Brontosaurus excelsus
"Noble thunder lizard"
Sobre esta espécie
Brontosaurus excelsus is one of the most iconic sauropods in the history of paleontology. It lived during the Late Jurassic, 156 to 146 million years ago, across the semi-arid floodplains of what is now the western United States. Up to 22 meters long and approximately 17 metric tons, it was one of the largest herbivores in its ecosystem. Its long, robust neck, far thicker than that of relatives like Diplodocus, may have served intraspecific combat roles in addition to feeding. Synonymized with Apatosaurus in 1903 by Elmer Riggs, the genus was revalidated in 2015 by Tschopp, Mateus, and Benson following the most comprehensive phylogenetic analysis ever conducted on diplodocids.
Geological formation & environment
The Morrison Formation is one of the most important sedimentary sequences for dinosaur paleontology in the world. Deposited between 156 and 147 million years ago during the Late Jurassic (Kimmeridgian to Tithonian ages), it covers much of the western United States, from New Mexico to Montana. The environment was a broad semi-arid alluvial plain with pronounced wet and dry seasons, crossed by seasonal rivers and dotted with shallow lakes. The formation preserved the greatest sauropod diversity known from a single ecosystem: more than ten genera, including Diplodocus, Brachiosaurus, Camarasaurus, Apatosaurus, and Brontosaurus excelsus itself.
Image gallery











Scientific reconstruction of Brontosaurus excelsus by Nobu Tamura (2016), showing the animal in lateral posture with robust neck and raised tail, consistent with the post-2015 scientific consensus.
Nobu Tamura — CC BY-SA 4.0
Ecology and behavior
Habitat
Brontosaurus excelsus inhabited the alluvial plains and conifer forests of the Morrison Formation, a semi-arid environment with distinct wet and dry seasons in what is now the western United States. The ecosystem was dominated by ferns, cycads, ginkgoes, horsetails, and conifers such as Brachyphyllum, with no flowering grasses. Seasonal rivers, shallow ponds, and floodplains provided water and concentrated the densest vegetation. It shared habitat with other giant sauropods including Diplodocus, Apatosaurus, Brachiosaurus, and Camarasaurus, as well as theropods like Allosaurus and Ceratosaurus and ornithischians like Stegosaurus.
Feeding
A strict herbivore, Brontosaurus excelsus fed on low to medium-height vegetation using its simple pencil-like teeth, concentrated at the front of a broad snout — an adaptation for nonselective, wide-sweep grazing. Dental microwear analyses in diplodocids suggest the animal would sweep vegetation with lateral head movements, consuming large volumes of ferns, horsetails, and low conifer foliage. Brontosaurus's exceptionally robust neck, distinct from the more gracile neck of Diplodocus, may have allowed access to denser or more resistant vegetation. Consumption estimates suggest an adult needed hundreds of kilograms of vegetation per day.
Behavior and senses
Behavioral evidence for Brontosaurus excelsus is inferred by comparison with relatives and through functional morphology. The exceptionally robust neck, with ventrally displaced cervical ribs and ventrolaterally directed parapophyseal rami, was interpreted by Taylor et al. (2015) as a possible adaptation for neck-to-neck combat between males, analogous to 'necking' behavior in giraffes. If confirmed, this hypothesis would imply complex social behavior with intraspecific competition for access to females. There is no direct evidence of herd behavior, but the abundance of specimens at some localities suggests individuals occupied common feeding areas.
Physiology and growth
Brontosaurus excelsus displayed the highly pneumatized axial skeleton typical of sauropods: vertebrae with internal cavities filled by air sac diverticula similar to the avian system, reducing neck and trunk weight without sacrificing structural strength. King et al. (2024) recently described pneumaticity also in the dorsal ribs — an unprecedented extension of the air sac system in apatosaurines. Bone histology indicates rapid, sustained growth in youth, suggesting elevated metabolism. Studies by Curry (1999) and Lehman & Woodward (2008) estimate the animal reached adult size in 15 to 70 years depending on the model. Thermoregulation likely benefited from gigantothermy: the enormous body volume retained metabolic heat even without full endothermy.
Paleogeography
Continental configuration

Ron Blakey · CC BY 3.0 · Jurassic, ~90 Ma
During the Kimmeridgiano–Titoniano (~156–146 Ma), Brontosaurus excelsus inhabited the fragmenting Pangea. North America and Europe were still close, and the North Atlantic was just beginning to open. Climate was warm and humid globally, with no polar ice caps.
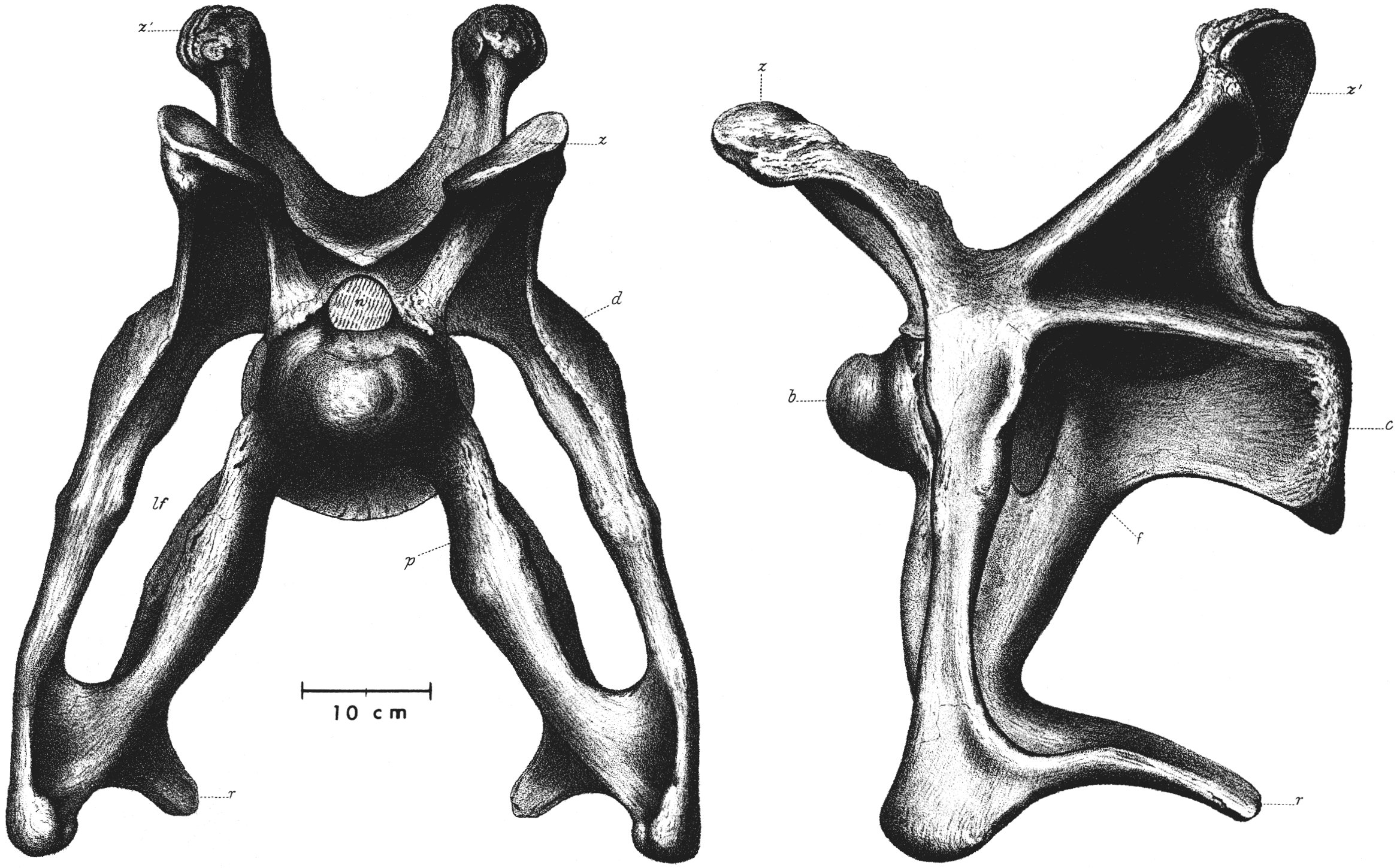
Inventário de Ossos
The holotype YPM 1980, collected at Como Bluff (Wyoming) in 1879, preserves vertebrae, ribs, pectoral girdle, and limbs, but lacked a skull. Specimen AMNH 460 (mounted in 1905 at the American Museum of Natural History) is a composite of several individuals. The true skull of the group was only established in 1975 by McIntosh and Berman, who demonstrated it resembles that of Diplodocus.
Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
Notice of New Jurassic Reptiles
Marsh, O.C. · American Journal of Science
The founding paper in which Othniel Charles Marsh names Brontosaurus excelsus based on specimen YPM 1980, collected by William Harlow Reed at Como Bluff, Wyoming. The description, published in the American Journal of Science in December 1879, establishes the genus based on cervical and dorsal vertebrae, ribs, pectoral girdle, and hind limbs. Marsh distinguishes Brontosaurus from Apatosaurus (described two years earlier) by differing vertebral and limb proportions, and notes the colossal size as exceptional. The name Brontosaurus — 'thunder lizard', from Greek brontē and sauros — and the epithet excelsus, from Latin for 'noble' or 'elevated', reflect the animal's perceived grandeur. This paper inaugurates one of the greatest scientific rivalries in history: the Bone Wars between Marsh and Edward Drinker Cope.
The Dinosaurs of North America
Marsh, O.C. · 16th Annual Report of the U.S. Geological Survey
Marsh's monumental work published as an annual report of the U.S. Geological Survey, presenting the first complete skeletal reconstruction of Brontosaurus excelsus. The illustration, based on holotype YPM 1980 supplemented by material from other Morrison Formation specimens, shows the animal in an upright posture with a slightly raised tail, defining the canonical sauropod image for the general public and science of the era. Marsh includes comparisons with other North American dinosaurs, discusses phylogenetic relationships of diplodocids, and proposes paleoecological interpretations of the Morrison Formation ecosystem. The work becomes an essential reference for all subsequent studies on Jurassic sauropodomorphs of North America.
Structure and Relationships of Opisthocoelian Dinosaurs. Part I, Apatosaurus Marsh
Riggs, E.S. · Publications of the Field Columbian Museum, Geological Series
Pivotal paper in which Elmer Riggs, from the Field Museum of Chicago, compares the Brontosaurus excelsus specimen from Como Bluff with a new Apatosaurus skeleton (P25112) collected from Fruita, Colorado. Riggs concludes that the diagnostic features Marsh used to separate Brontosaurus from Apatosaurus — vertebral and limb proportions — actually reflect ontogenetic differences, with the Apatosaurus ajax holotype being an immature individual. Thus, by the principle of nomenclatural priority, Apatosaurus (1877) takes precedence over Brontosaurus (1879), and Riggs creates the combination Apatosaurus excelsus. Published in a relatively obscure journal, Riggs' argument took decades to be fully absorbed by the scientific community and the public.
Description of the Palate and Lower Jaw of the Sauropod Dinosaur Diplodocus (Reptilia: Saurischia) with Remarks on the Nature of the Skull of Apatosaurus
McIntosh, J.S. & Berman, D.S. · Journal of Paleontology
Revolutionary study correcting a 70-year error in Brontosaurus/Apatosaurus reconstruction. Since 1905, museum mounts had displayed Camarasaurus-type skulls — robust, with spatulate teeth — because no skull had been found with postcranial skeletons. McIntosh and Berman analyze available skull specimens and demonstrate that the correct cranial morphology is diplodocid: narrow, low skull with simple pencil-like teeth concentrated at the front of the mouth. The discovery of a skull associated with specimen CM 3018 (Carnegie Museum) in 1909, which had remained unstudied for decades, confirms the conclusion. The correction is adopted by the Carnegie Museum in 1979, when the first correct skull is mounted on an apatosaurine skeleton — four years after this paper's publication.
Supersonic Sauropods? Tail Dynamics in the Diplodocids
Myhrvold, N.P. & Currie, P.J. · Paleobiology
Pioneering study by Nathan Myhrvold and Philip Currie using computer simulation to model the tail dynamics of Apatosaurus louisae — a taxon closely related to Brontosaurus excelsus. Results suggest the long, tapering diplodocid tail could be cracked like a bullwhip, with the tip reaching supersonic velocities and producing a sonic boom analogous to a real whip crack. The hypothesis proposes this behavior served communicative or defensive functions. Published in Paleobiology in 1997, the paper generated enormous scientific and media impact, though later analyses contested whether supersonic velocities were truly achievable given muscular friction and air resistance. Even if partially refuted, the paper established a new paradigm in sauropod paleoethology.
Ontogenetic histology of Apatosaurus (Dinosauria: Sauropoda): new insights on growth rates and longevity
Curry, K.A. · Journal of Vertebrate Paleontology
Classic histological study by Kristina Curry analyzing bone microstructure in an ontogenetic series of Apatosaurus radii, ulnae, and scapulae — a genus directly related to Brontosaurus excelsus within Apatosaurinae. Examination of growth lines (LAGs) and fibrolamellar bone tissue reveals three distinct osteogenic phases: rapid sustained growth during most of ontogeny, gradual deceleration approaching adult size, and eventual cessation. Age estimates of ~10 years for large sub-adults refute the hypothesis that slow, indeterminate growth was needed for sauropods to achieve extreme sizes. The paper inaugurated systematic application of bone histology to sauropods and established the 'bird-like' rapid growth paradigm for the group.
Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs
Stevens, K.A. & Parrish, J.M. · Science
Seminal study by Kent Stevens and Michael Parrish using articulated digital reconstructions of Apatosaurus and Diplodocus necks — taxa fundamental to understanding Brontosaurus excelsus ecology — to infer neutral cervical posture. Using the DinoMorph program, the authors model intervertebral joints and conclude both diplodocids held their necks in a slightly downward-inclined position at rest, with the head near ground level. The result suggests low-level feeding on ferns and low vegetation, not high-canopy browsing. The paper generated intense debate: Taylor et al. (2009) would argue that neutral posture in living terrestrial animals corresponds to maximum sustainable elevation, implying raised necks. The controversy shaped two decades of sauropod paleoecology research.

Modeling growth rates for sauropod dinosaurs
Lehman, T.M. & Woodward, H.N. · Paleobiology
Lehman and Woodward analyze histological sections of Apatosaurus long bones — directly related to Brontosaurus excelsus — combining growth line (LAG) counts with body mass estimates to model growth curves. Two alternative models are tested: one based on bone length-to-mass relationships suggesting adult size (~25,000 kg) reached in as few as 15 years at a maximum rate of 5,000 kg/yr; and a von Bertalanffy model projecting 70 years to adult size at a maximum rate of 520 kg/yr. The discrepancy reflects fundamental uncertainties in the relationship between histology, mass, and metabolism in Jurassic giants. The paper is a central reference in the debate over whether sauropods were endothermic, ectothermic, or mesothermic.

Why sauropods had long necks; and why giraffes have short necks
Taylor, M.P. & Wedel, M.J. · PeerJ
Taylor and Wedel analyze why sauropod necks — including Brontosaurus excelsus, which possessed a notably robust neck for a diplodocid — reached lengths of up to 15 meters, six times longer than any giraffe record. The paper systematically evaluates six hypotheses: access to high-canopy vegetation, feeding on aquatic surfaces, sweeping a wide area without moving the body, thermoregulation, sexual selection, and neutral evolution. The conclusion is that feeding advantage — exclusive access to plant resources beyond the reach of other herbivores — is the primary selective factor, with sexual selection as a plausible secondary factor.


Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses
Whitlock, J.A. · PLOS ONE
Whitlock combines snout shape analysis and dental microwear microscopy to infer feeding behavior in diplodocoids, the group including Brontosaurus excelsus. The study identifies two distinct feeding strategies: diplodocids with squared snouts engaged in nonselective, wide-gape feeding sweeping broad areas at ground level, while brachiosaur relatives with rounder, more robust snouts engaged in selective browsing at greater height. Brontosaurus/Apatosaurus, with its intermediate snout and exceptionally robust neck, appears to have occupied an ecologically distinct niche from other family members. The paper provides the first quantitative microwear-based evidence for dietary niche partitioning among Morrison Formation's giant herbivores.


Were the necks of Apatosaurus and Brontosaurus adapted for combat?
Taylor, M.P., Wedel, M.J., Naish, D. & Engh, B. · PeerJ PrePrints
Taylor, Wedel, Naish, and Engh examine morphological features of the Apatosaurus and Brontosaurus neck — specifically ventral displacement of cervical ribs and ventrolaterally directed parapophyseal rami — arguing these structures are consistent with adaptations for intraspecific neck-to-neck combat, analogous to 'necking' observed in giraffes. Brontosaurus's exceptionally robust, muscular neck, in contrast to Diplodocus's more gracile neck, is seen as evidence that different diplodocid genera used their necks for different functions. The combat hypothesis would imply Brontosaurus had more complex social behavior than previously assumed, with males competing for access to females or resources.


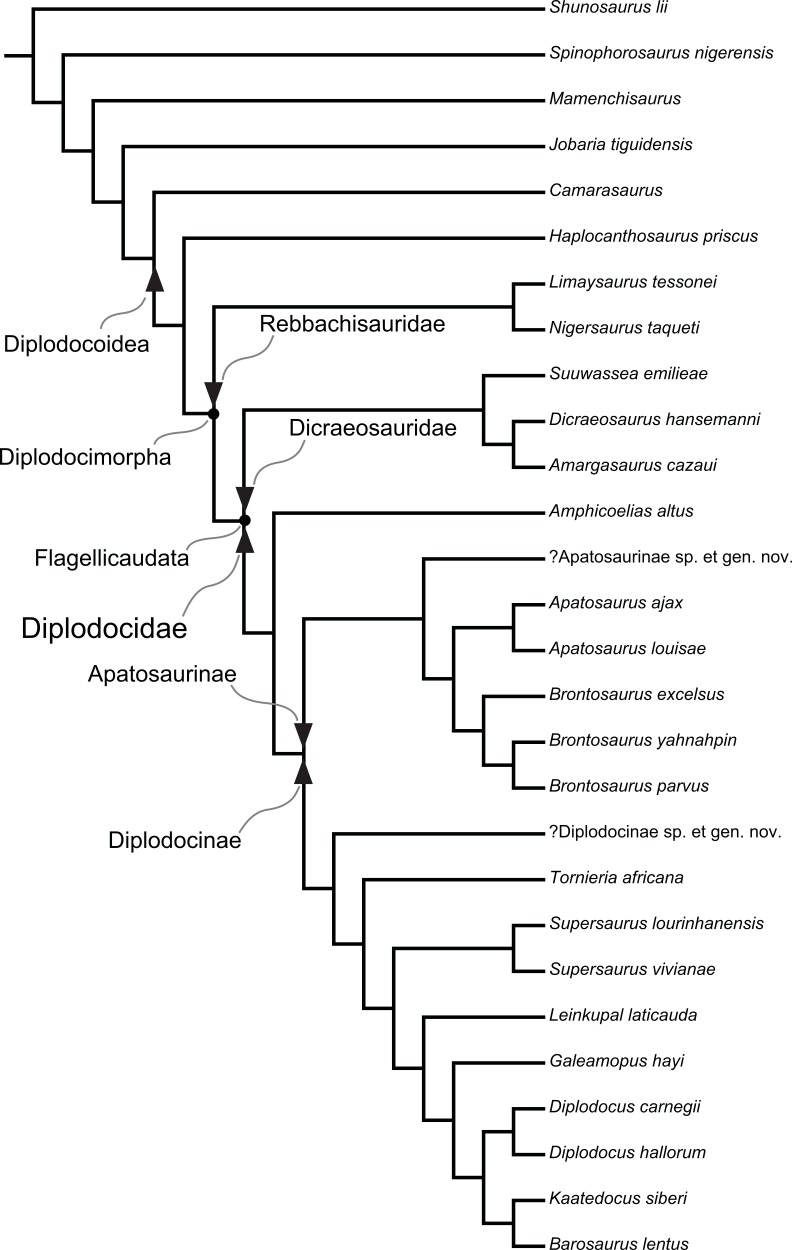
A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)
Tschopp, E., Mateus, O. & Benson, R.B.J. · PeerJ
The Brontosaurus revalidation paper, published in PeerJ in 2015, is the most comprehensive phylogenetic work ever conducted on Diplodocidae. Tschopp, Mateus, and Benson score 477 morphological characters across 81 sauropod specimens — a five-year analysis involving visits to museums in Europe and the United States. The innovative specimen-level approach (rather than species-level) allows detection of subtle differences between historically confused taxa. The result is unequivocal: Brontosaurus presents sufficient autapomorphies (unique characteristics) distinguishing it from Apatosaurus, revalidating the genus after 112 years of synonymy. Three species are recognized: B. excelsus (type), B. parvus, and B. yahnahpin. The paper generated global media coverage.


Evidence for niche partitioning among ground-height browsing sauropods from the Upper Jurassic Morrison Formation of North America
McHugh, J.B. · Geology of the Intermountain West
McHugh examines isotopic and morphometric evidence from Morrison Formation sauropods to investigate how multiple giant genera — including Brontosaurus excelsus — coexisted in the same ecosystem without direct food competition. The study focuses on ground-height browsers, proposing that differences in tooth morphology, snout shape, and neck movement range allowed each genus to exploit distinct feeding microhabitats. Brontosaurus, with its exceptionally robust and muscular neck, may have exploited dense or resistant vegetation inaccessible to other diplodocids. The open-access paper contributes to resolving the 'Morrison diversity paradox' — the extraordinary coexistence of more than ten sauropod genera in a semi-arid environment with limited food resources.


A previously unreported form of dorsal rib pneumaticity in Apatosaurus and Brontosaurus
King, A., Wedel, M.J. & Taylor, M.P. · PeerJ
King, Wedel, and Taylor describe, for the first time, pneumaticity in the dorsal ribs of Apatosaurus and Brontosaurus — a type of internal air cavity similar to those found in birds, extended beyond the vertebrae (already known as pneumatized) into the ribs. The study uses CT scanning and surface examination of specimens in museum collections to document pneumatic chambers and fossae within costal bone tissue. The discovery implies that the air sac system — fundamental to efficient pulmonary ventilation in birds — was even more extensive in apatosaurines than previously thought, with diverticula penetrating the axial skeleton more broadly than recognized.


Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs
Wedel, M.J. · Paleobiology
Wedel analyzes vertebral pneumaticity in sauropods — a system of internal chambers and fossae in vertebrae filled by air sac diverticula similar to the modern avian system. The study documents the phylogenetic distribution of pneumaticity in sauropods: basal taxa only in presacral vertebrae, derived forms (like diplodocids, including Brontosaurus excelsus) with extension into anterior caudals. The presence of avian-style air sacs implies a unidirectional airflow respiratory system — far more efficient than the bidirectional mammalian lung — consistent with high metabolic rates. The paper established vertebral pneumaticity as a fundamental diagnostic and physiological tool in sauropod paleobiology, and serves as the reference work preceding King et al.'s (2024) discovery of rib pneumaticity in Brontosaurus.


Espécimes famosos em museus
YPM 1980 (Holótipo)
Yale Peabody Museum of Natural History, New Haven, Connecticut, EUA
Type specimen of the species, collected at Como Bluff, Wyoming. Preserves cervical and dorsal vertebrae, ribs, pectoral girdle, and fore and hind limbs, but no skull. On permanent display in the Peabody Museum's Great Hall since 1931, mounted at 20 meters in length.
AMNH 460 (Monte Composto)
American Museum of Natural History, Nova York, EUA
Historic mount inaugurated in 1905 as the world's first sauropod skeleton. Composed mainly of specimen AMNH 460 from Nine Mile Crossing Quarry (Como Bluff), supplemented by bones from AMNH 592, 222, and 339 and plaster casts of holotype YPM 1980. Tschopp et al. (2015) classified this specimen as 'indeterminate apatosaurine' pending detailed analysis.
In cinema and popular culture
Brontosaurus is perhaps the dinosaur most present in the cultural imagination of the 20th century. The name entered popular vocabulary even before being synonymized with Apatosaurus in 1903 — and remained a synonym for 'long-necked dinosaur' in the public mind for over a century, regardless of any taxonomic revision. In cinema, the trajectory begins in 1925 with 'The Lost World', where a Brontosaurus is the central character of the London climax, and continues in 1933 with 'King Kong', where the animal appears as a fearsome predator on Skull Island. In 1940, Disney immortalized it in 'Fantasia' grazing in Jurassic landscapes to the sound of Stravinsky. Decades later, in 1993, 'Jurassic Park' mentioned the name as a child's confusion — a precise reflection of how Brontosaurus lived in popular vocabulary even when it was scientifically synonymous with Apatosaurus. In 2015, Pixar's 'The Good Dinosaur' featured an apatosaurine protagonist the same year Tschopp et al. revalidated the genus. The 2015 scientific revalidation closed a 112-year cycle: Brontosaurus became a valid name again — and pop culture, which never abandoned it, can finally say it was right all along.

Classificação

Descoberta
Curiosidade
Brontosaurus went extinct twice: first as a living animal (146 million years ago) and then as a valid scientific name — suppressed for 112 years (1903–2015) until being 'resurrected' by the most comprehensive phylogenetic study ever conducted on diplodocids. It is the only dinosaur in history to have its genus officially abolished and then restored.