Scientific Repository
Dinos101
Explore each species discovered by paleontology: where it lived, which bones were found, and how our understanding evolved over time.
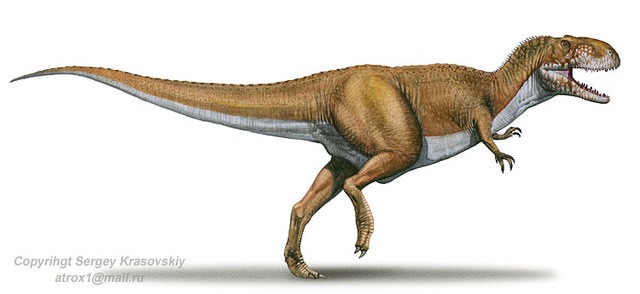

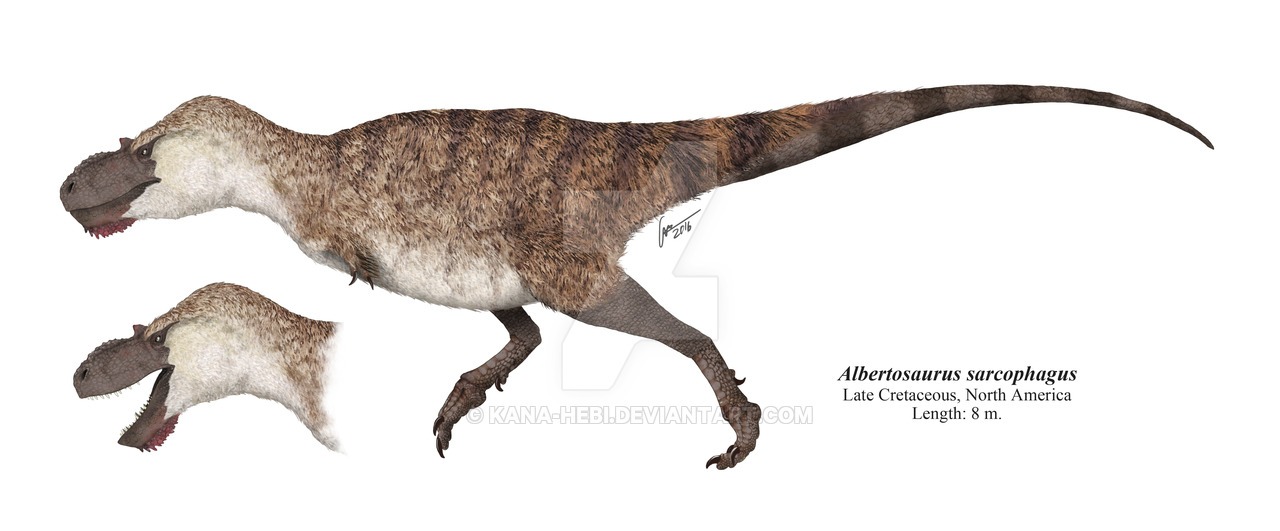



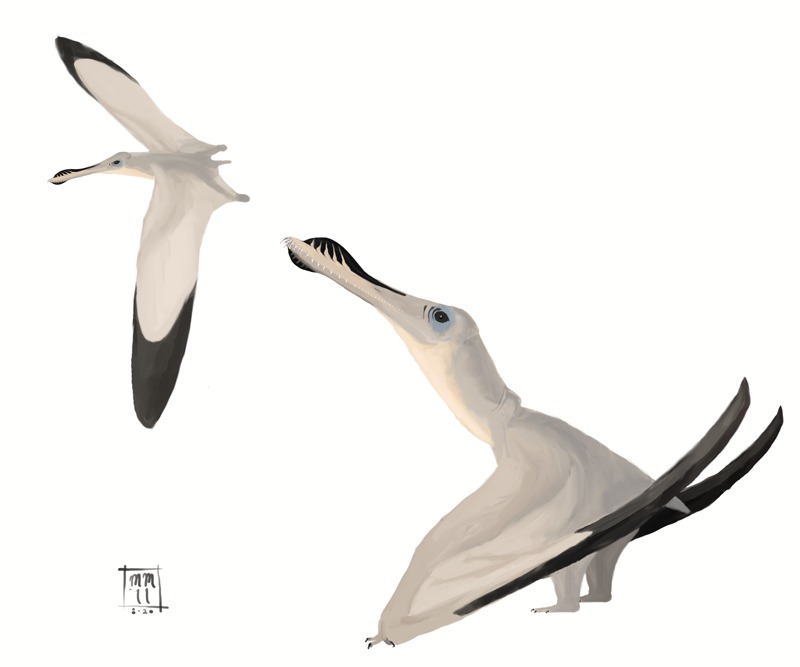



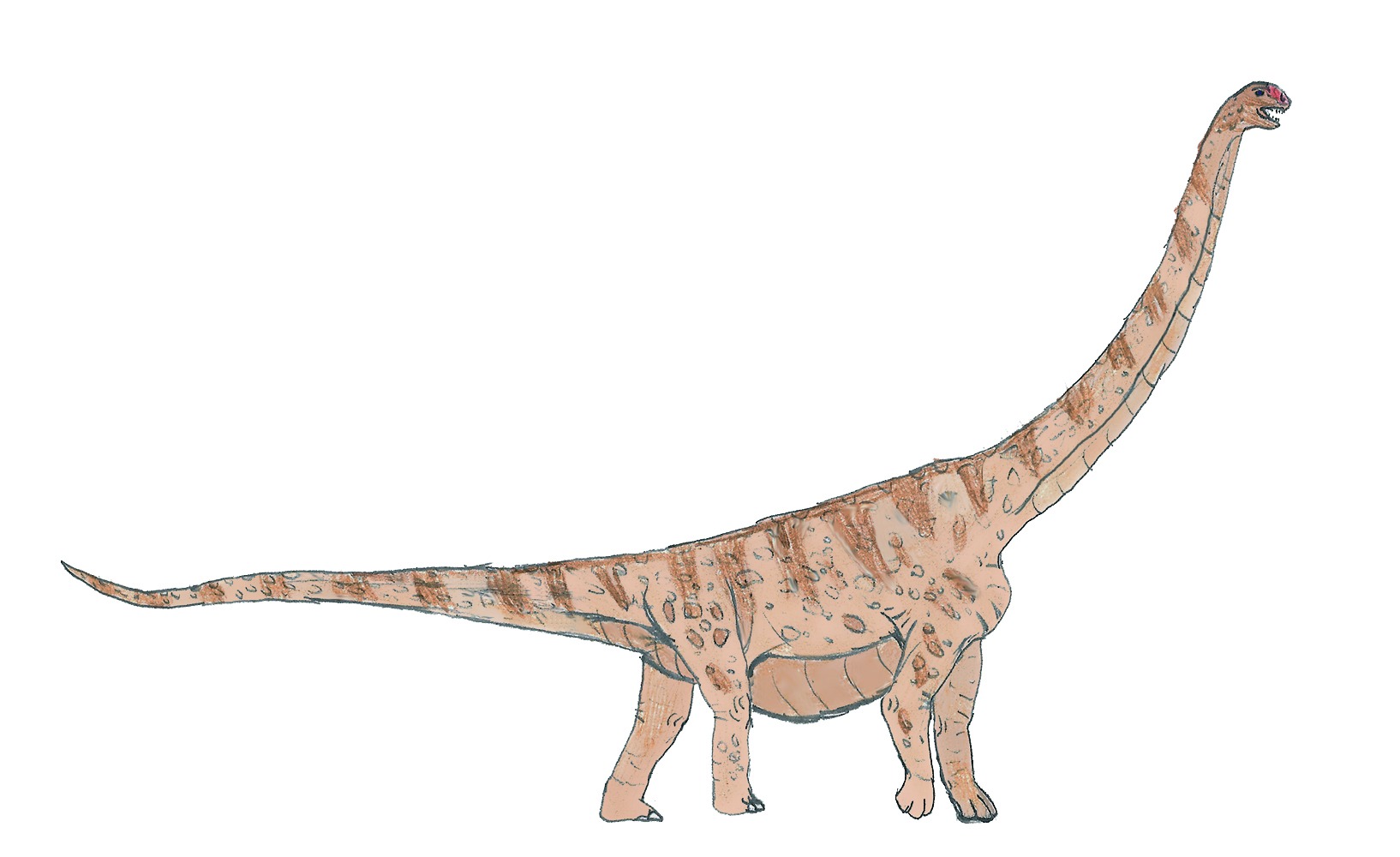









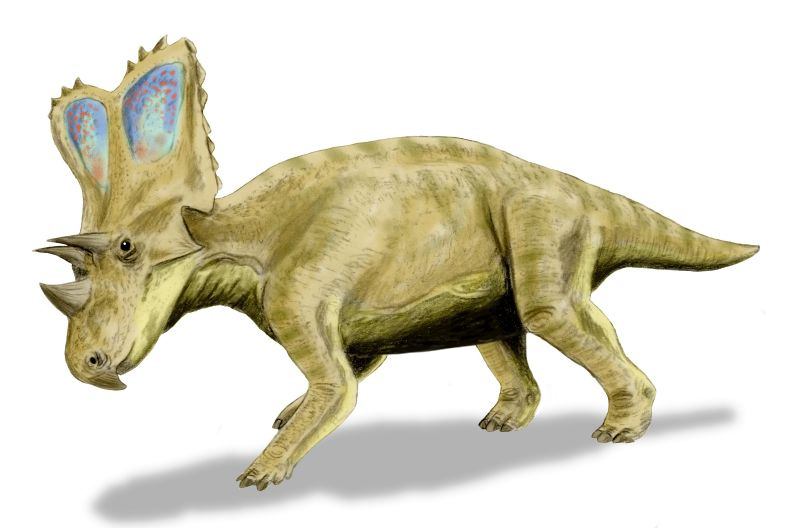


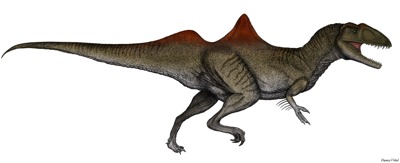


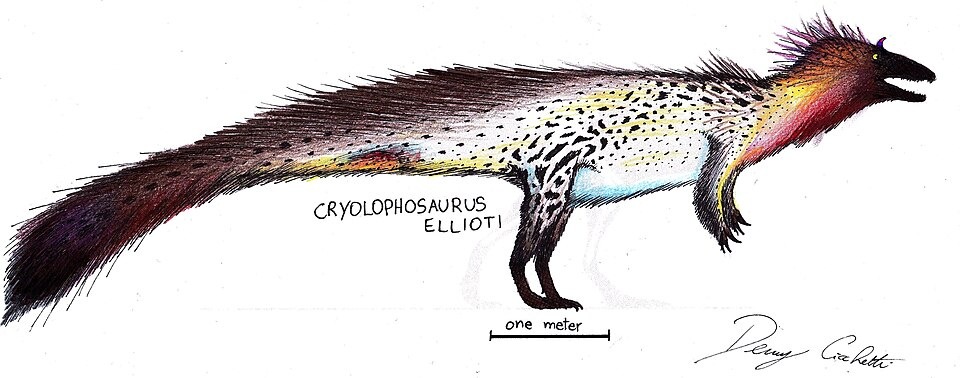
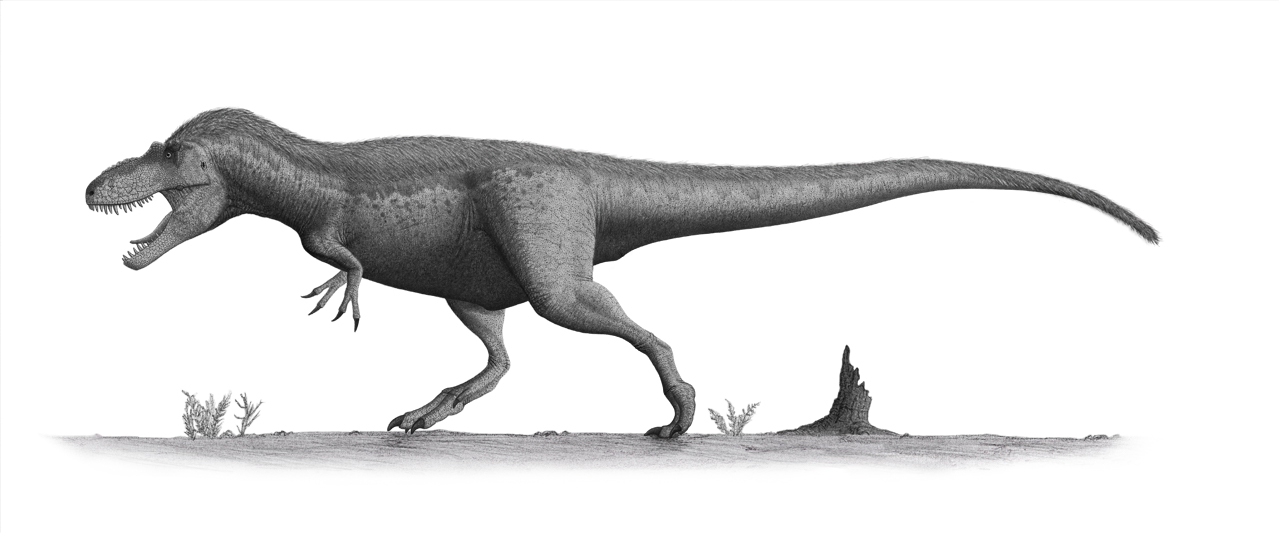


















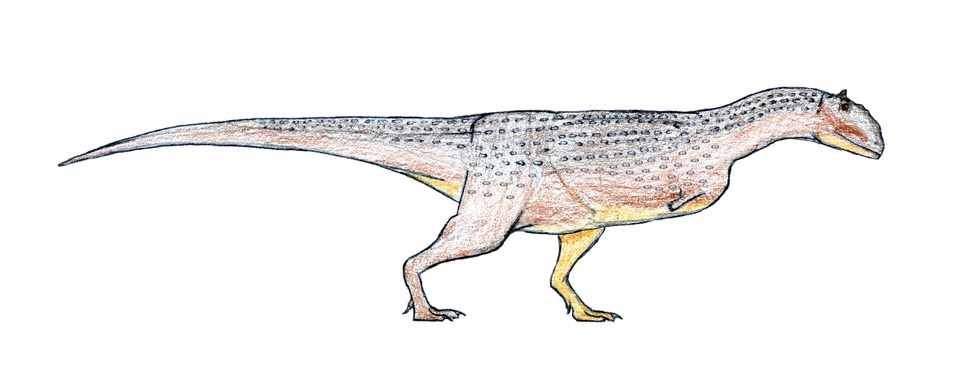


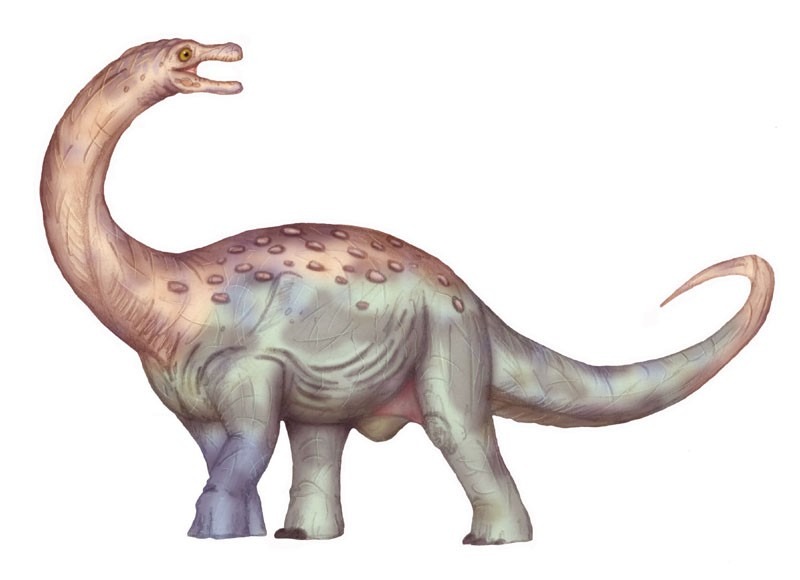


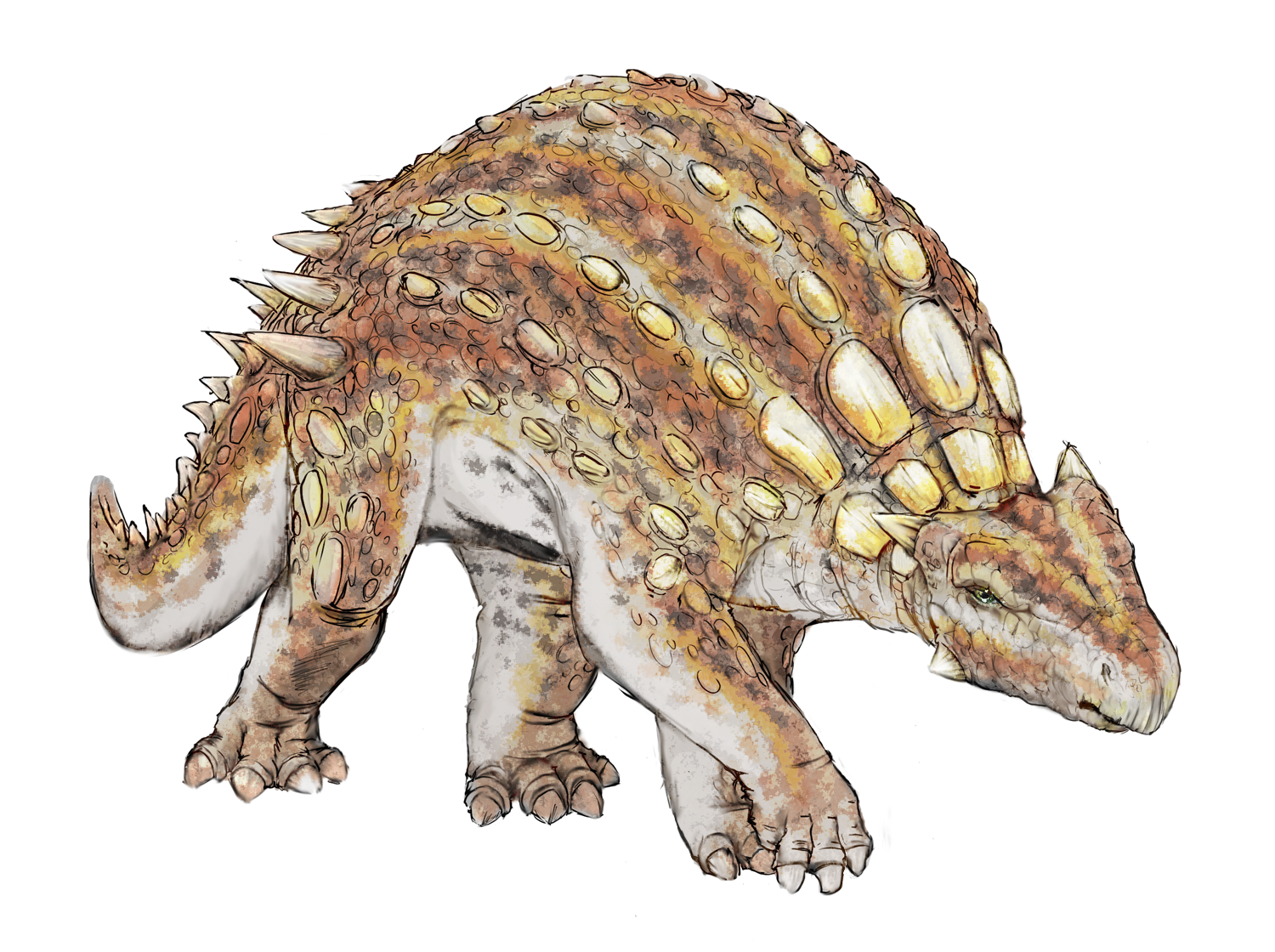
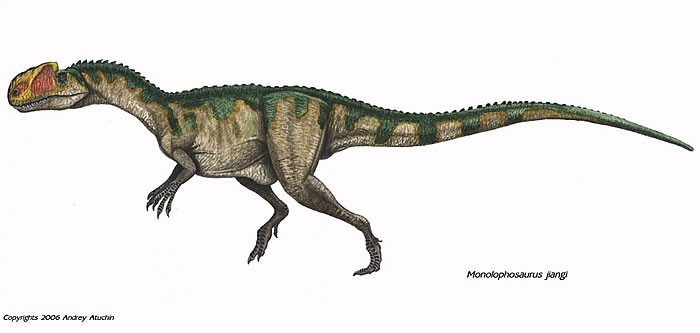


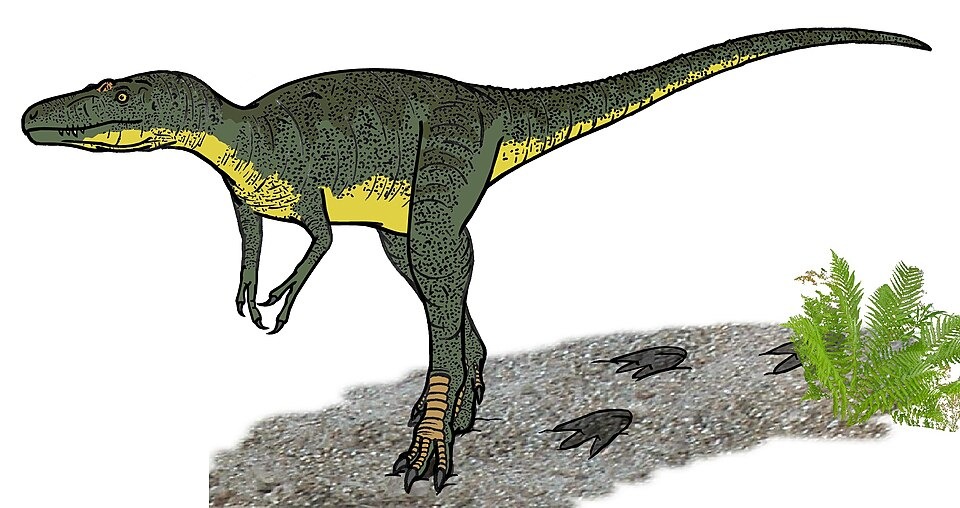
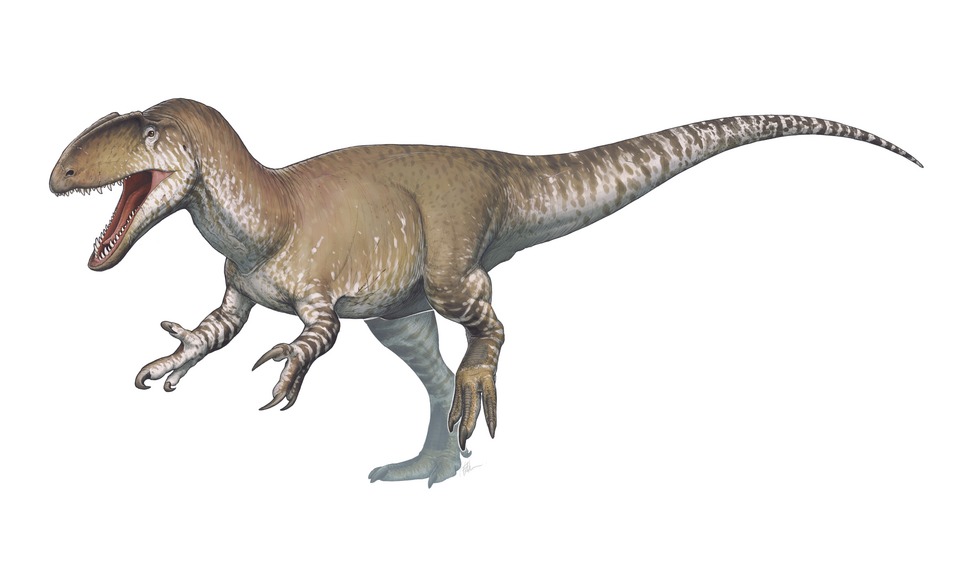
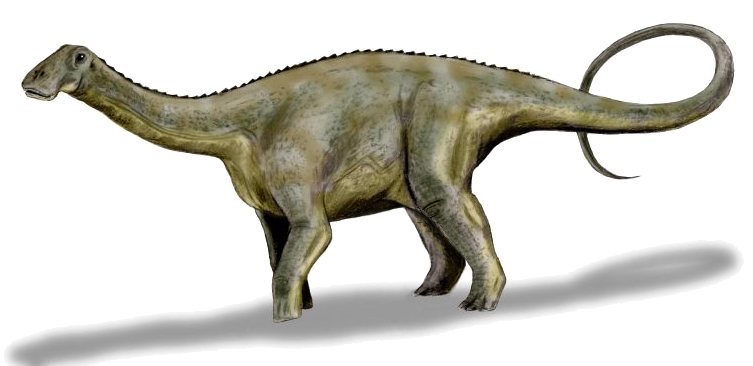








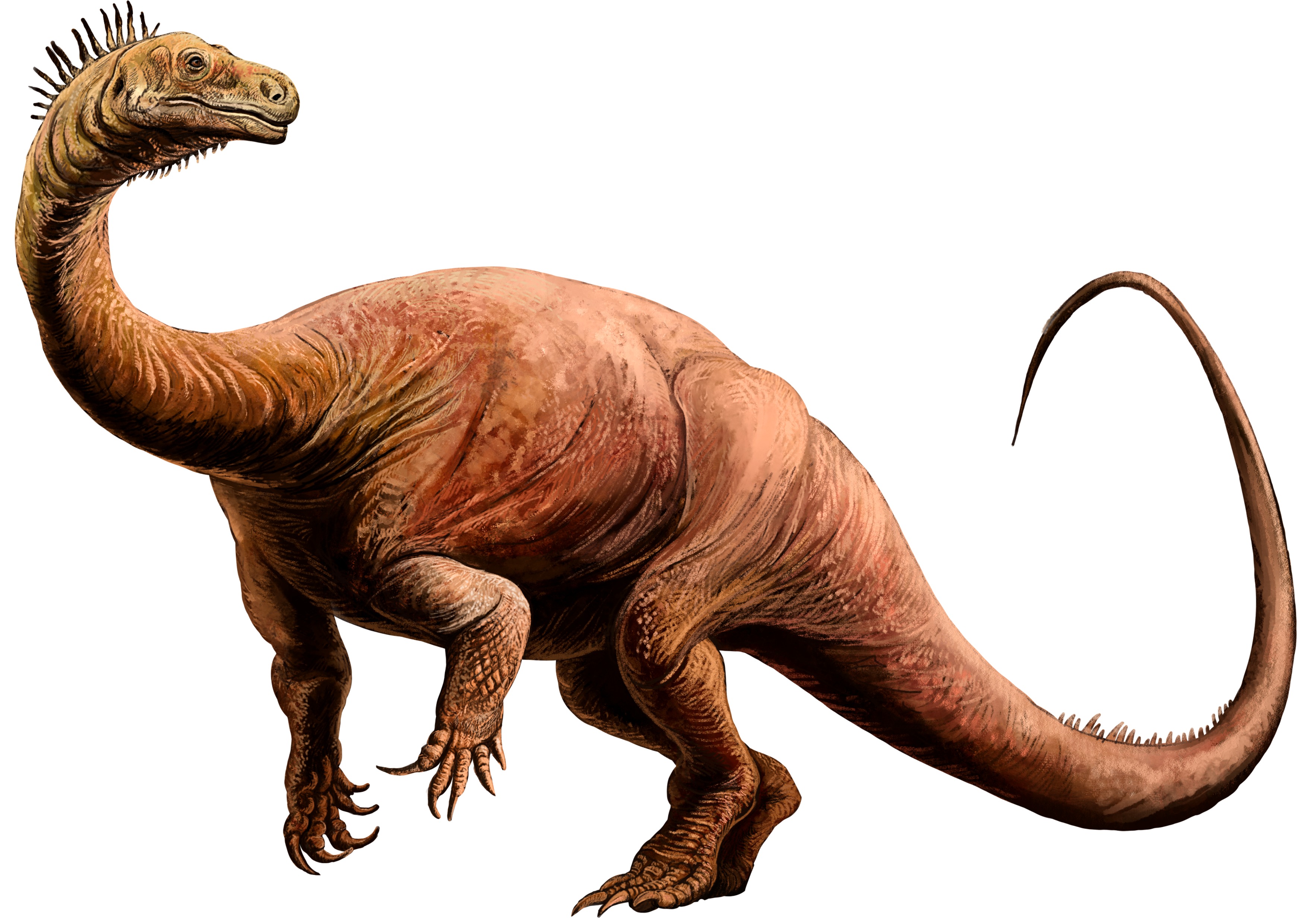




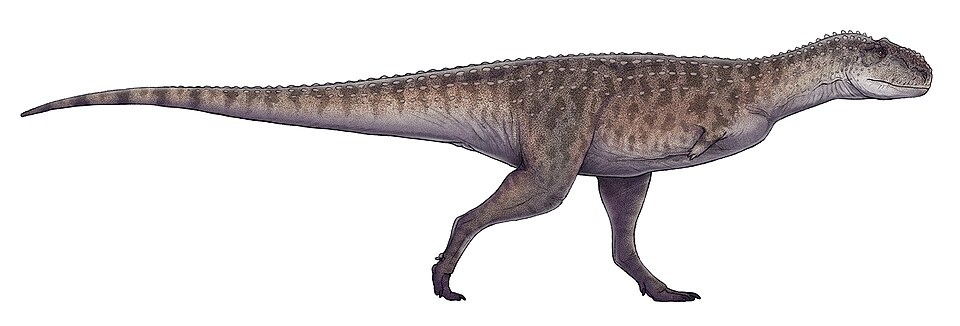
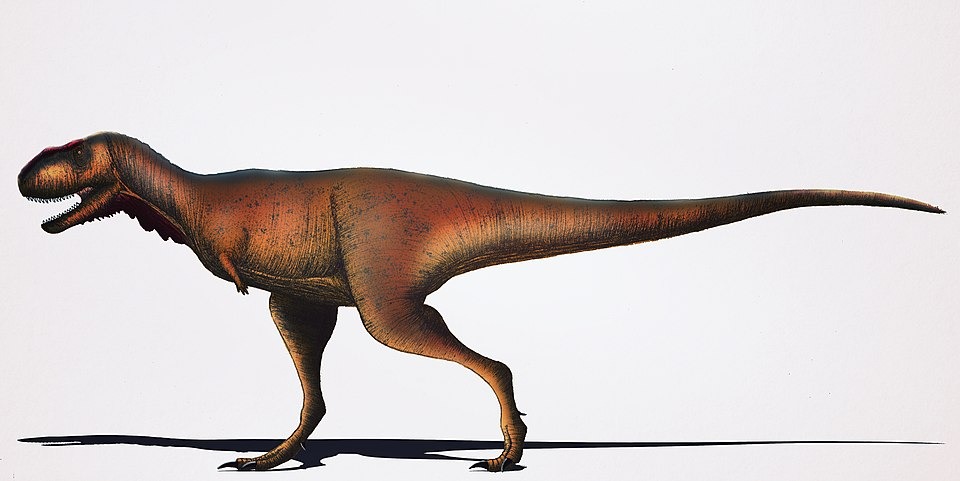




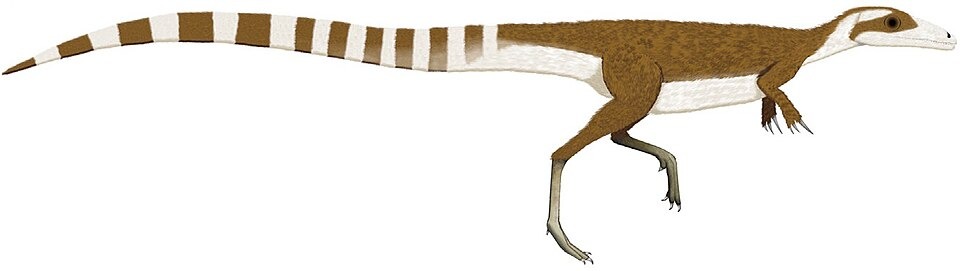
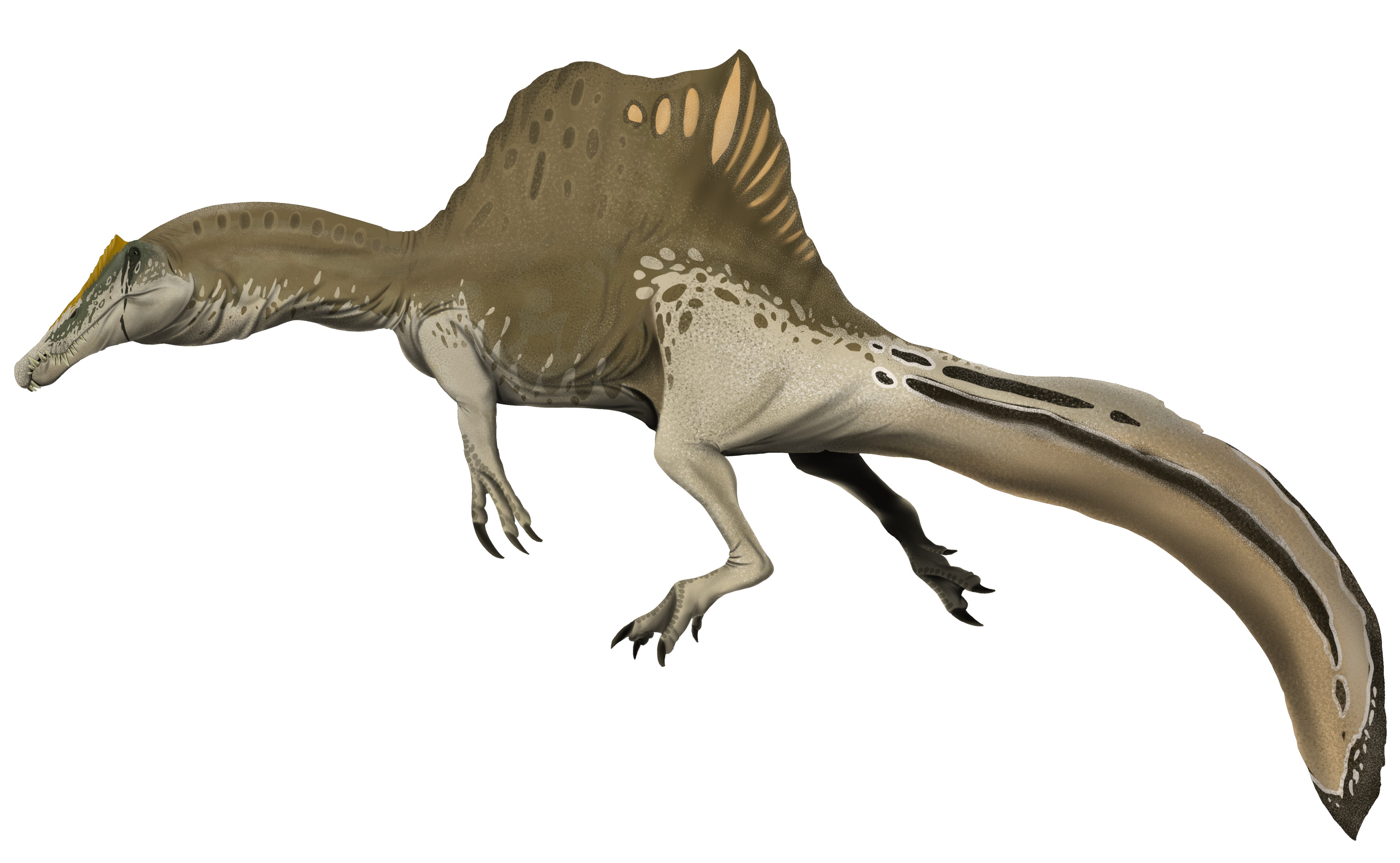
















No species found with these filters.
Mesozoic · Non-dinosaurs
Other Animals of the Mesozoic
Not dinosaurs, but they dominated the oceans, the skies, and parts of the continents during the same period.

Shark · Marine
Hybodus
"Curved tooth"
The most successful shark of the Mesozoic. Had two types of teeth: pointed for fish and flat for crustaceans — rare versatility. Shared the seas with ichthyosaurs, plesiosaurs, and mosasaurs.

Shark · Marine
Cretoxyrhina mantelli
"Sharp-snouted of the Cretaceous"
One of the largest sharks of the Cretaceous. Smooth unserrated teeth, high speed, morphology similar to the modern great white. Fossils show mosasaur vertebrae bearing its tooth marks.

Shark · Marine
Squalicorax
"Raven shark"
Shark with heavily serrated teeth similar to the modern tiger shark. Fossil evidence shows it fed on carcasses of dinosaurs that fell into rivers and seas — the open-ocean scavenger niche.

Ichthyosaur · Marine
Ichthyosaurs
"Fish lizards"
Extraordinary convergence with dolphins and sharks, but they were reptiles that gave birth to live young at sea. Shonisaurus (Triassic) reached 21m. Ophthalmosaurus had the largest proportional eyes of any known vertebrate.

Plesiosaur · Marine
Plesiosaurs
"Near lizard"
Elasmosaurus had ~14m of neck with 72 cervical vertebrae. They swam with four large flippers in a kind of "underwater flight," snatching individual fish with rapid neck strikes.

Plesiosaur · Marine
Pliosaurs
"More lizard"
Sister group of plesiosaurs, but with a short neck and enormous head. Kronosaurus had a 2.7m skull. Predator X (Jurassic, Norway) may have reached 15m. Top predators of Jurassic oceans.
Mosasaur · Marine
Mosasaurus hoffmannii
"Lizard of the Meuse River"
The largest known mosasaur at 17 meters. Related to monitor lizards and snakes, not crocodiles. First giant marine reptile fossil described by science (1764, Maastricht). Had a double-hinged jaw like snakes.

Pterosaur · Aerial
Pterosaurs
"Winged lizards"
Not dinosaurs — they were the sister group. Had hair-like fibers (pycnofibers) and were likely endothermic. Quetzalcoatlus northropi: 10–11m wingspan, 2.5m tall on land. Largest flying animal of all time.
Archosaur · Terrestrial
Rauisuchids
"Crocodilians of Rau"
While dinosaurs were still small, rauisuchids were the apex predators of the Triassic. Saurosuchus (7m), Postosuchus (6m), Fasolasuchus (8–10m). Their extinction cleared the way for large theropods.

Archosaur · Terrestrial
Phytosaurs
"Plant lizards"
Perfect convergence with modern crocodilians, but no close relation. Key difference: nostrils positioned almost between the eyes (in crocodilians they sit at the tip of the snout).

Crocodylomorph · Terrestrial
Crocodyliformes
"Crocodile-shaped"
Far more diverse in the Mesozoic than today: bipedal terrestrial forms in the Triassic, pelagic marine forms in the Jurassic. Dakosaurus had a theropod-like skull. What we know today is just the survivors.

Synapsid · Terrestrial
Dicynodonts
"Two dog teeth"
Synapsids with a horny beak and tusks. Lystrosaurus dominated the early Triassic after surviving the Permian mass extinction. Placerias (hippo-sized) briefly coexisted with the first dinosaurs.

Mammal · Terrestrial
Mesozoic mammals
"Mammals of the Age of Dinosaurs"
Small, likely nocturnal, living on the margins. But there was real diversity: Castorocauda (semi-aquatic, 164 Ma), Volaticotherium (glider, 160 Ma), Repenomamus (fossil found with a dinosaur hatchling in its stomach).